We've been looking at the process of
starting currents using generator potentials.
Introduction
We've been
looking at how our bio-electrical system works.
A current
is passed along the wires (excitable cells) of our
bioelectrical systems.
Charge (a
current) is the only thing that moves in an electrical
circuit.
The current
is pushed by a voltage.
Bioelectrical
systems have such a high internal resistance that the
voltage to keep the current moving must be continually
recreated along the path of the current.
The
physiological event that recreates the voltage along the
pathway is called an action potential.
The
triggering event for an action potential is the increase
in membrane potential increase caused by the rapidly
dissipating currrent created directly upstream from the
site of the AP.
We call
this membrane potential increase a generator potential.
An AP
is always caused by a generator potential.
In our
sensory system, the first generator potential is created by
a receptor cell.
Every
sensory receptor cell has solved a transduction problem.
The
transduction problem that was solved was how to create a
current that is proportional in magnitude to the
particular phenomenon being sensed (sound, touch, taste,
smell, etc.)
The
receptor cell therefore creates the first generator
potential along the sensory pathway.
In some
cases, the solution is to distort the receptor cell or
appendages on the receptor cell to mechanically open ion
channels that create the generator potential (pressure,
hearing, balance).
In some
cases, the solution is to use a sensing protein that is
sensitive (changes its structure in response) to the
environmental phenomenon.
This
structural change is detected by an intracellular
signalling system (e.g., the G-protein system) that will
then activate a ligand gated ion channel that creates
the generator potential.
The
ligand attachment site is in the cytosol, rather than
the extracellular fluid (as was the case for transfer of
the current across a synapse).
Examples:
vision, smell (olfaction), some tastes (e.g., sweet).
The current
that is passed from the sensory receptor cell to the brain
is periodically broken by the end of one neuron and the
beginning of another.
A synapse
consists of the terminal bulb-gap-section of the
dendrite/cell body that handles all the steps of
recreating the current on the other side of the gap
between the cells.
(the
synaptic cleft is the gap between the cells).
The
details of the process were handled in earlier readings.
The
magnitude of the generator potential created in the
receiving neuron determines whether the current is passed
along the next neuron.
The
magnitude of the generator potential from a single
terminal bulb can be augmented by the next generator
potential from the same terminal bulb (temporal
summation).
The
frequency of the AP's in the preceeding neuron must be
great enough to support rapid sequential synaptic
transmissions.
OR
multiple terminal bulbs from the same neuron must be
involved (spatial summation).
In either
case, the neurotransmitting substance must be excitatory
(resulting in opening of ligand gated Na channels on the
down-side of the synapse).
It is
also possible to suppress current transmission.
Terminal
bulbs are either excitatory OR inhibitory (they cannot
be both, they cannot change function).
Terminal
bulbs from a different neuron may prevent the passage of
a current (or reduce it's magnitude) through the use of
inhibitory neurotransmitting substances.
Inhibiting
NTS activates ligand gated potassium or ligand gated
choride channels to reduce the magnitude of the
generator potential by decreasing the positive charge
concentration.
Creating a
generator potential in the electrical pathway is the first event
in the transport of a current along afferent neurons (those
carrying the current to the brain/CNS) and efferent neurons
(those carrying the current from the brain/CNS to the peripheral
parts of the body.
The afferent
neurons that carry information to the brain are hard wired
into the parts of the brain that will initiate the processing
of the information.
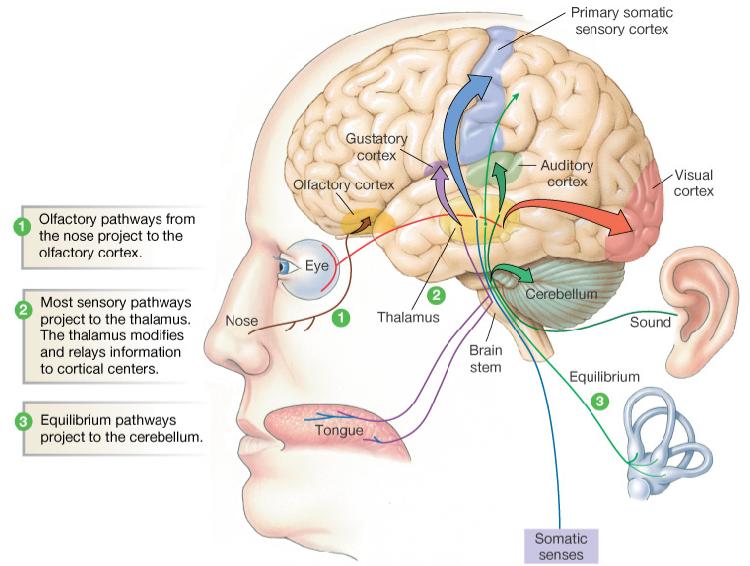
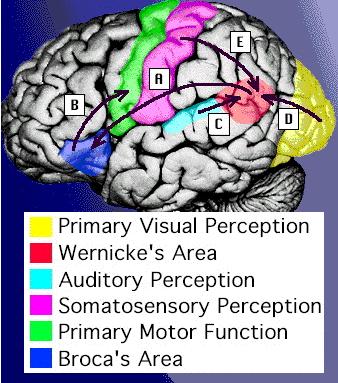
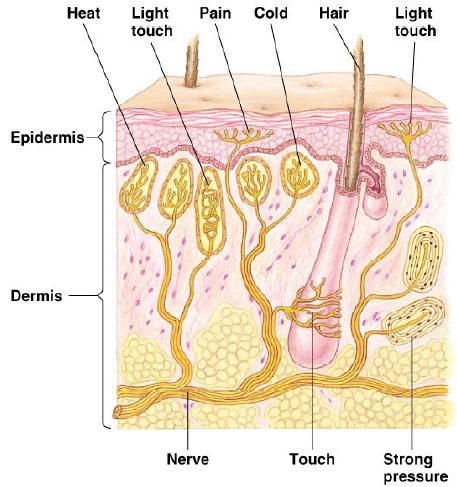
Somatic
senses: (touch, pressure, heat, cold, and pain)
are hard wired into the brain.
These
are are passed up the brain stem.
All
but pain are routed to the somatic sensory
cortex located along a strip across the top
surface of the brain.
Pain
signals communicate with the cortex and with
other parts of the brain.
All
the other sensory inputs have centers located in
different parts of the brain.
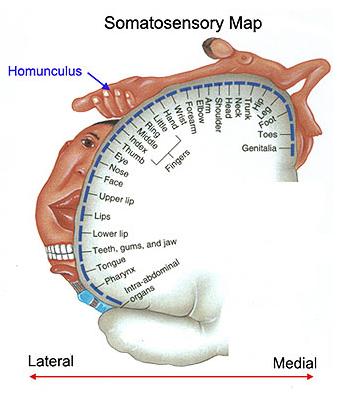
The
figure at right shows the somatic sensory cortex on
the left side of the brain.
The
sensory cortex in the left brain receives
information from the somatic neurons from the
right side of the body, and vice-versa.
As
illustrated by the figure, somatic inputs are not
evenly distributed along the body surface:
Face,
tongue, hands, and genitals have greater somatic
sensitivity than arms, legs, back, etc.
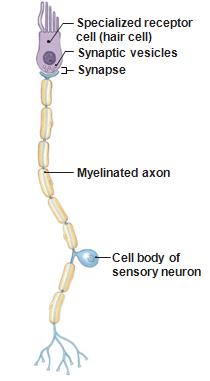
Sensory
receptors are sometimes stand alone cells and sometimes
modified dendrites connected to a much larger neuron.
Notice
the nucleus in the hair cell.
The
receptor is a fully functional cell not
dependent upon the sensory neuron for control,
resources, etc.
Taste
and vision receptors were similar.
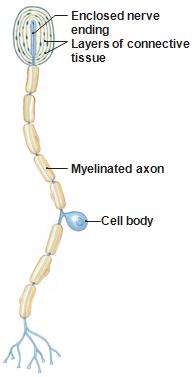
For
other receptors, the sensory apparatus is mounted
on modified dendrites of the neuron that also
carries the signal.
Pacinian
corpuscles and olfactory neurons are like this.
Many
of the pain receptors that we will talk about
today are also "modified nerve endings".
Repeating
something ...
In addition to
solving the transduction problem, sensory recptors have solved
a much more basic problem.
They have solved
the problem of how to create a current in our electrical
system.
We have the
wires.
Wires don't
produce currents.
Where do
the currents that pass through the wires come from?
Now we know
about the currents that tell us about our environment.
What about
the current that constitutes a decision to act or a thought
(that originates in the brain)? Where do these currents come
from?
A Bit More on
Vision
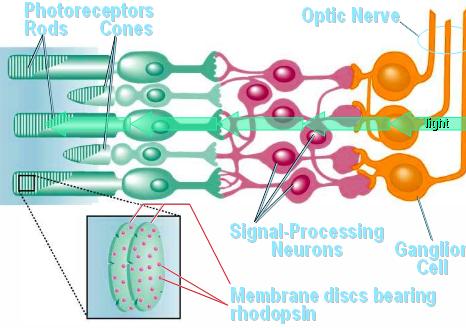
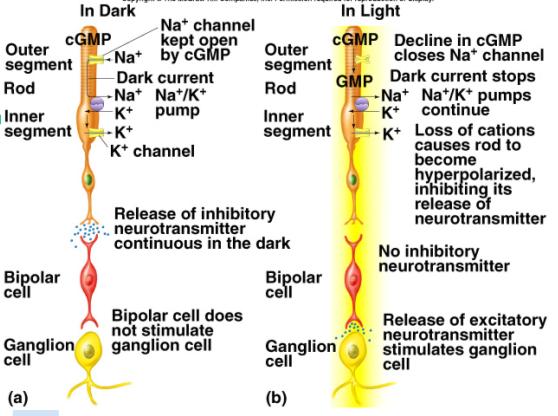
Rods and cones
are photo receptors. The other receptors respond to chemicals
(chemoreceptors) or mechanical events (streching, movement of
stereo cilia, etc., mechanoreceptors)
You saw
last time that light must pass through the ganglion
and bipolar cells before it reaches the light
sensitive receptor cells.
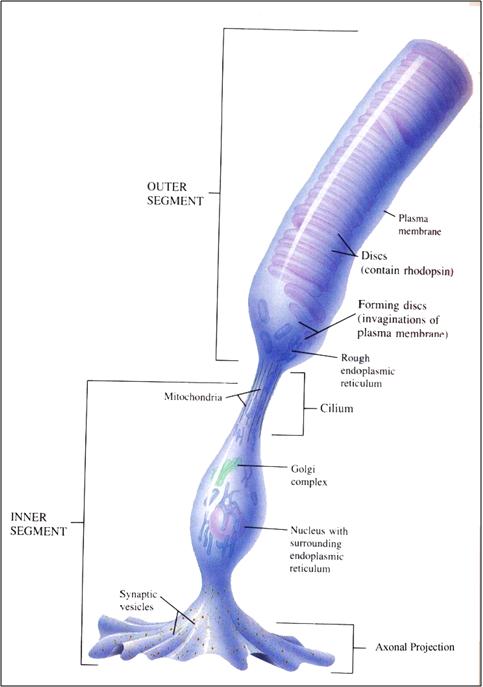
The
pigment that responds to photons is located within
the rod or cone.
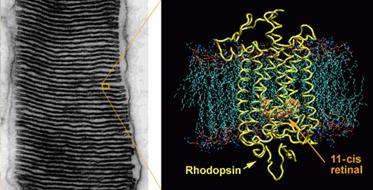
Here is
a closeup showing the arrangement of pigment discs
within a rod.
On
the left is another view of the disks in a
rod.
On
the right is the protein rhodopsin in the
structural mesh that makes up the disc.
Rhodopsin
contains the 11-cis retinol that allows vision to
occur.
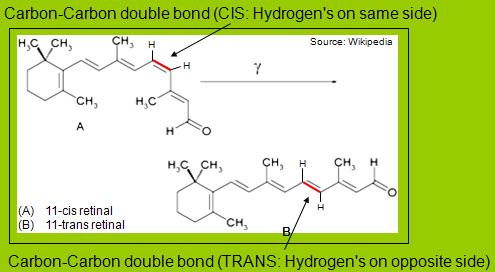
When a
photon strikes the retinol, it causes a C-C bond to
break and the cis-retinol changes its conformation
(into the trans form).
The
retinol is embedded within the protein rhodopsin.
What
do you think happens to the rhodpsin when the
retinol changes conformation?
What
do you think this initiates?
Just for review: here is the sequence
of events that we dicussed last time.
More
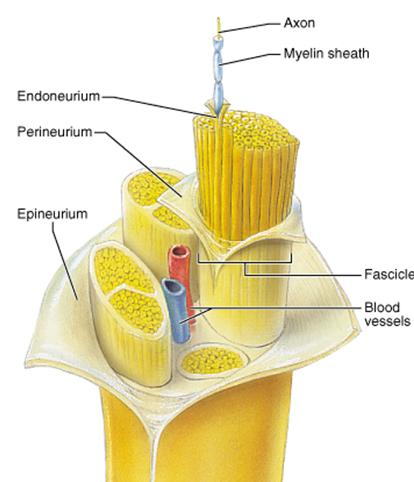
Review: Neural Superhighways. In an earlier
reading, we referred to aggregates of neurons that are
bundled together in the central nervous system.
recall
that their myelin came from oligodendricytes
rather than Schwann cells.
Note
that bundles of neurons (each having it's own
covering) are called fascicles.
You
will see this term again when we get to skeletal
muscles.
Courtesy
J.R.
Schiller, Austin Peay State University
Pain
We haven't
talked much about the brain yet. We'll start to do so (a bit)
today.
Pain is
fundamentally different than our other sensations.
Not in the
methods used to generate currents.
Pain is
pretty conventional, in that sense.
Rather, it
differs in what it is looking at and where it goes when it
arrives in the brain and How it is processed.
In all of
the other senses, we are solving the transduction problem
for a specific environmental event or process.
Salt in
your mouth.
Photon
intensity, density, and wavelength in your eyes.
Pressure
waves in your ear.
With pain,
we are looking for artifacts of damage.
Burning
Intense
cold
Ripping
of tissue
Intense
pressure
Events
related to disease.
All of
these relate to specific types of damage.
Also,
for sight, hearing, touch, smell, and taste, there
are specific regions of the brain to which the
neurons for that particular sense are "hard
wired".
In the
brain, however, the currents produced by pain receptors
do not go to a specific "center".
Rather,
their path and processing is more diffuse,
creating
an emotional response and
interacting
with cognitive (higher order, reasoning) functions
in the cerebral cortex.
There
is much about pain at this end that is not well
understood.
Flinching
dogs.
Long term
debilitating effects of torture.
The
anticipation of the syringe is usually worse than the fact.
This is a
very different response than we have to other electrical
inputs through our afferent sensory neurons.
Even at the
receptor cell level, there are some interesting difference.
Last
video (does a good summing up before we look at
specifics).
Pain Receptor
Cells Pain receptors do not fall into a neat package.
In fact, it's such a hodge-podge that if it wasn't so
important I would skip it. But it is important and we will
step through some of the basic information on how the pain
transduction problem works.
For most of our
senses, it's just a transduction problem, and we are aware of
the specific entity that we sense:
Photons in
the visible range - sight
Compressional
waves in air - hearing
Chemicals
floating around in the air - smell
Pressure on
our skin - touch
Chemicals
dissolved in our saliva - taste
This is not the
case with pain. We don't say:
"The pH has
dropped in my finger." Instead, we say ouch!
"A cell in
my finger has split open." Instead, we say ouch!
"The
potassium concentration of the extracellular fluid in my
finger has increased." Instead, we say ouch!
In general, what we
perceive as pain is induced by chemical changes that we wouldn't
necessarily associate with pain.
But this is what
the sensors that perceive pain (the "nociceptors") respond to.
The Role of
Pain
Pain
differs from other systems in that it is a "damage alert"
system.
Vision,
smell, and hearing may alert you that a predator is
approaching.
The pain
of the attack alerts you that the predator has inflicted
damage.
Purpose of
pain?
Avoidance
of additional damage (eat on the other side of your
mouth).
Allow
healing to occur (rest).
What is Pain?
In the
Transport class, you should have learned that there is no
such thing as heat.
There is
thermal kinetic energy that our temperature receptor cells
respond to and we perceive as "heat" or "cold".
Similarly,
there is no such thing as pain.
There are
pain receptor cells (called nociceptors) that respond to a
variety of environmental events.
When they
create a current in response to one or more of these
events, we perceive it as pain in the pain centers of the
brain.
There is
also nueropathic pain (e.g., referred pain, phantom
pain, pinched nerves) that is a currents in a pain
conducting neuron that is initiated somewhere (other than
the receptor) along the elctrical neural pathway.
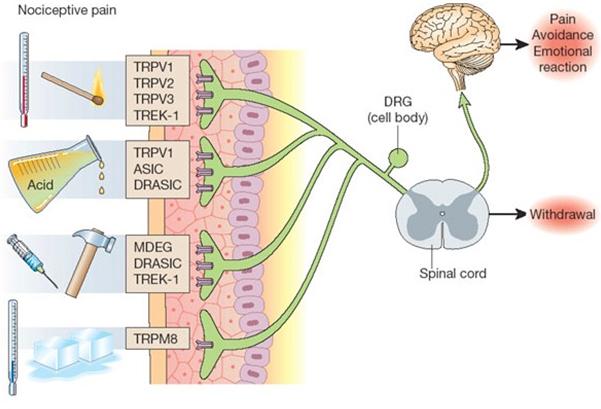
Pain
receptors are found in many parts of the body (skin,
muscles, joints, some internal organs
There
are 5 general types of nociceptors:
Thermal
- Detect hot or cold noxious stimuli.
Different
than receptors that respond to a little bit warm
or cool.
Mechanical
- Detect noxious pressure or deformation, such an
an incision.
Are
activated only by extreme deformation (e.g.,
stretching or pressure).
Chemical
- Respond to many different types of
chemicals.
Polymodal
"Sleeping"
or "silent" - Only respond to post-injury
inflammation.
Many
sensory receptors are just modified dendrites of a
generalized neural cell (they resemble pacinian
corpuscles, in that respect).
The
modification consists of the actual receptor sites
located on neural dendrites.
We
saw this appraoch (different chemical receptors on
otherwise identical cells) when we discussed
olfaction (smell).
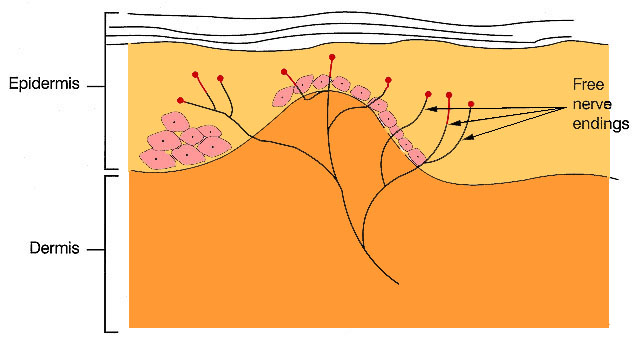
Nociceptors
are the undifferentiated terminals of small myelinated
or unmyelinated neurons.
Pain Receptors. There are a number of
receptors involved in transduction of the pain stimuli into a
current carried to the brain.
Heat
sensitive receptors. Above a threshold of 43 C these
heat activated channels will open, allow cations (Na+
in particular) to flow into the cell, thus causing a
depolarization. The greater the stimulus temperature above
threshold the greater the current (the greater the frequency
of action potentials). These neurons are distinct from the
heat sensitive sensory neurons which will respond to
increasing temperatures that are non-painful. The two types of
sensory neurons are active in different temperature ranges).
Receptors
associated with tissue damage. A number of
chemoreceptors respond to chemical changes associated with
tissue damage.
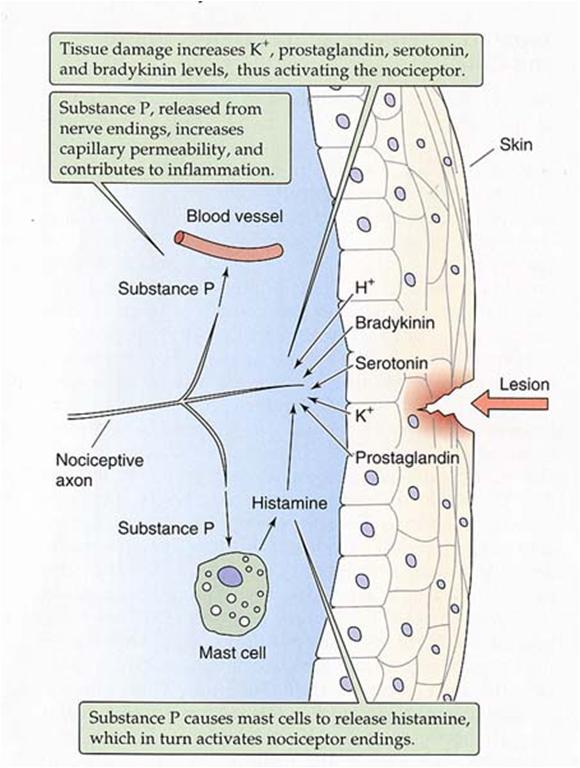
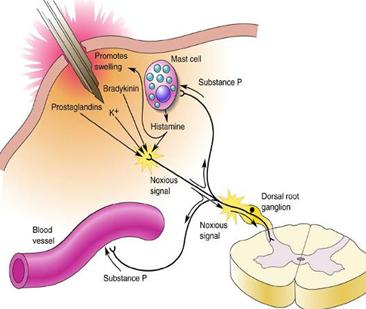
After a
significant injury several things happen in the vicinity of
the damage:
There
is a local decrease in pH as the contents
of cells spills into the tissue, as cell interiors
are generally more acid than the extracellular
space.
There
is an outflow of potassium ions from
ruptured cell contents.
There
is an outflow of ATP from ruptured cell
contents.
Leakage
of plasma occurs from damaged capillaries.
The
leaking plasma contains a variety of chemicals
and enzymes which influence the local tissue
cells.
The
most important for pain production is
bradykinin.
Receptor types
that respond to these chemical changes include:
pH-gated channels. These
are H+ sensing channels that responds to pH below
6.5 .
These
receptors are also stimulated by exercise (a carbon
dioxide increase associated with exercise lowers pH in the
vicinity of the exercising muscle).
This
appears to be the same mechanism as capsaicin receptor
(taste that detects spiciness of chili peppers).
Both Na
and Ca flow through same channels and contribute to the
generator potential.
ATP
gated channels. ATP is released from damaged cells.
ATP-gated ion channels are found in nociceptors (ATP is the
ligand in these ligand gated channels).
Bradykinin.
Damaged cells release proteolytic enzymes which result in
the break down of a preprotein into a peptide called
bradykinin.
Bradykinin
binds to a receptor that activates a G protein sequence.
The G
protein sequence includes a second messenger that acts as
the ligand for a ligand gated Na+ channel to
depolarize the sensory neuron (as seen in olfaction)
Other substances
increase sensitivity of nociceptors (make them more liable to
create currents):
Potassium
leakage from ruptured cells.
Potassium
increases membrane potential without opening channels.
Increasing
[K]out decreases the negative current
(leakiness of the cell membrane to potassium) and raises
the nernst potential.
This
results in increased membrane potential (as described by
the Goldman equation).
Serotonin.
Released from platelets (platelets are cell fragments that
originate in bone marrow and are essential for clotting).
Histamine.
Released from mast cells (mast cells are what control many
of the body's allergic reactions). When the body comes into
contact with an allergen, the mast cells release
histamine-containing granules. A series of events unfold,
ultimately causing swelling.
Other chemicals produced when cell enzymes enter
the extracellular fluid (i.e., the plasma) include
prostaglandins.
Prostaglandins are created when cellular enzymes
act on lipid
precursors.
The prostaglandins initiate inflammation.
Athough not
specifically mentioned in the
video at right, prostaglandins
vasodilate arterioles and
increase permeability of
capillaries.
The video
does a nice job of
describing acute inflammation (which
includes redness, increased
temperature, swelling, pain, and
increased sensitivity of
nociceptors).
Notice
the reference to
histamine production
caused by wounds
(which mediates
vasodilation)
and serotonin which acts
on nociceptors as another
pain inducing
agent.
We can understand
vasodilation
as a relaxing
of the smooth
muscles that
surround
arterioles and
some venules.
How can we
understand
increased
permeability
of
capillaries?
substance
P - released from active nociceptors - provides
additional excitation along the neural pathway (summation)
to make the pain more "noticable".
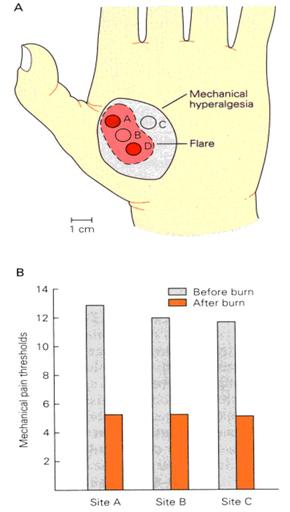
Sensitization can
extend to areas away from the original damage, resulting in
hypersensitivity.
The mode of
action in hypersensitivity is still under investigation:
In some
cases, receptor cells implicated in hypersensitivity create
a cross membrane curent which increases membrane potential,
moving it closer to the threshold required for a generator
potential to initiate a current to the brain.
In this
case, a smaller generator potential in the nociceptor
results in a current to the CNS indicating a painful
experience.
In other
cases, it appears that the chemicals alter the protein
conformation of the receptor, directly increasing the
receptor's sensitivity to the noxious stimulus.
In this
case, the receptor responds at a lower level of
stimulation, producing a generator potential that leads to
an AP with a smaller provocation (e.g., a very light
bump).
Desensitization can
also occur, for example when non-steroidal anti-inflammatory
analgesic drugs including aspirin inhibit the production of
prostaglandin.
Visceral Pain
"Visceral pain
is pain that results from the activation of nociceptors of the
thoracic, pelvic, or abdominal viscera (organs).
Visceral
structures are highly sensitive to distension (stretch),
ischemia (loss of oxygen supply), and inflammation.
They are
relatively insensitive to other stimuli that normally evoke
pain such as cutting or burning.
Visceral
pain is diffuse, difficult to localize and often referred
to a distant, usually superficial, structure.
It may be
accompanied by symptoms such as nausea, vomiting, changes in
vital signs as well as emotional manifestations.
The pain
may be described as sickening, deep, squeezing, and dull."
(paraphrased from Wikipedia).
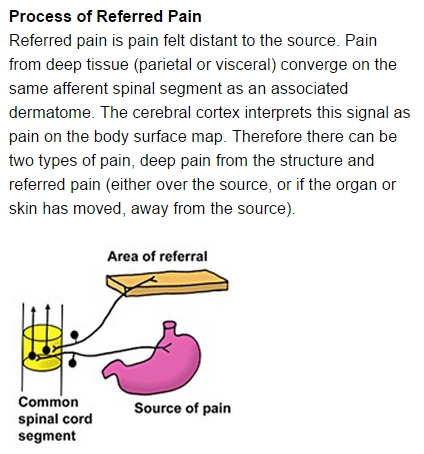
Referred Pain
The presumption
that pain originates at the perceived site of that pain can be
incorrect. Sometimes the neural signal that tells us that a
particular part of the body "hurts" does not originate at a
receptor cell but rather along the path between the receptor
cell and the brain. This is called "referred" pain.
Referred pain is
pain from a malfunctioning or diseased area of the body that
is perceived as though it was originating in another area,
often far from the actual site of damage or disease. A common
example is found in a person having a heart attack. In this
case, pain may be experienced down the inside of the left arm
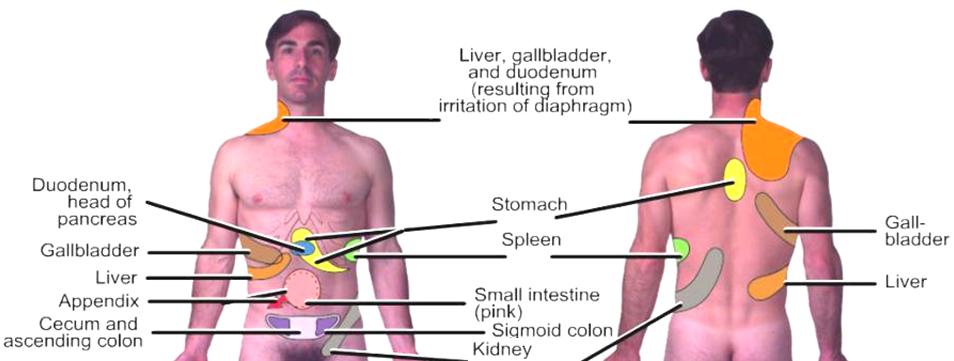
and forearm. There are other common manifistations, including:
the gall
bladder referring pain on the top of the right shoulder.
a diaphragm
problem may be felt in the shoulder and neck.
stomach
problems may refer to the spine between the shoulder blades.
kidney pain
may be felt in the groin area.
a problem
in the throat may be referred to the ear.
intestinal
dysfunction may be felt in the middle or low back.
Pain from
visceral organs and other interior structures is transported
via type C (unmyelinated) fibers. In many cases these are
transported along common pathways with type A (myelinated)
neurons that service surface nociceptors.
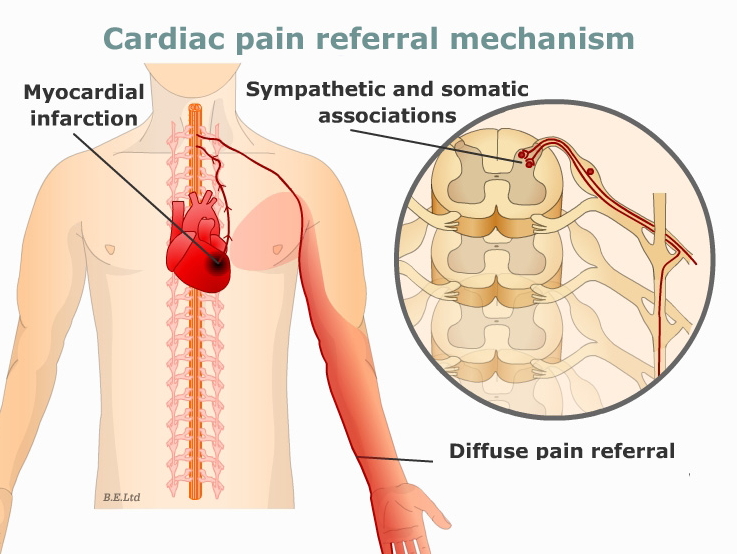
In
the case of referred pain associated with heart
problems, it is known that pain from angina or
myocardial infarction (the term for an actual heart
attack is "myocardial infarction") is transported
via afferent nerve fibers that enter the spinal cord
on the left side (see figure at right).
In
the process of entering the spinal cord, the
afferent neuron from the heart passes close to the
primary and secondary somatic neurons that
communicate pain from the left arm.
The
actual process of how a current in one neuron creates a
current in an adjacent neuron is not completely
understood.
The
conventional wisdom, at present, is that the
unmyelinated afferent pain neuron connecting the
organ to the spinal column passes near to the cell
body or dendrites of a secondary afferent neural
pathway that services another part of the body (the
"area of referral" in the figure at right).
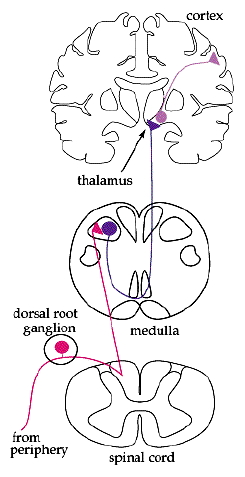
A
complete neural pathway consists of multiple
neurons (see figure, far right),
For
receptor pathways, the primary neuron carries
the current from the receptor to the spinal
cord.
A
secondary neuron then carries the current to the
brain (usually the thalamus) where it is passed
off to a tertiary neuron that transports the
current within the brain (usually to a location
in the cerebral cortex).
The
local electrical activity along the unmyelinated
deep organ path appears to create a generator
potential in the secondary neuron that services the
area of referral.
Since
the axon of the primary neuron that services the
arm is myelinated, it is less likely that the
induced generator potential would occur there.
It
is more likely to induce the generator potential
in a dendrite or the cell body of the secondary
neuron.
In
this way, a current is created that appears to
indicate that pain is coming from the area of referral.
In
the case of a myocardial infarction (heart attack)
the referral area is the left arm.
For other
internal organs the same phenomenon involves other referral
areas. These are shown, below.
How Does a
Local Anesthetic Work? (source: Stan Lee-Son)
Local
anesthetics block conduction along nerve pathways by
inhibiting the creation of action potentials.
During
depolarization, the major excitatory process is the opening
of sodium channels to allow Na+ ions into the neuron.
If you
can block depolarization, you will block or reduce the
propagation of the current to the part of the brain that
processes pain.
A
local anesthetic binds to sodium channels,
blocking Na+ transport during
depolarization.
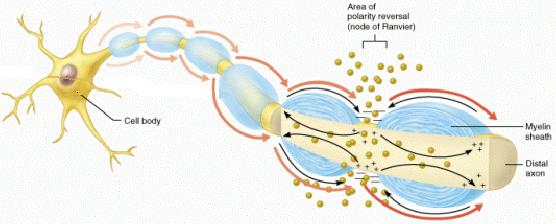
In
myelinated neurons, it is assumed that 3
consecutive nodes of Ranvier must be effected to
halt action potential propagation.
This
is partly because it is difficult to block all
of the sodium channels at any one node of
Ranvier under physiological conditions.
It
is also because a very strong receptor potential
(generator potential created by the receptor)
may be strong enough to propagate a current to
the next location where an AP occurs, if the
first location fails to depolarize.
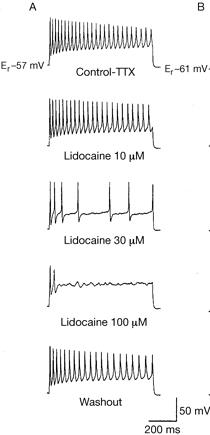
The
figure at right shows the response of a
myelinated rat neuron to increased concentrations of
lidocaine.
The
neuron was electrically stimulated.
Dosing
was adjusted to create the lidacaine
concentrations at
The
measurements were taken at a site beyond the
effected nodes of Ranvier.
Notice
the attenuated response at 30 uM and the complete
loss of activity at 100 uM.
At
30 uM, the primary observable response
downstream from the effected nodes is a
reduction in the frequency of the measured AP's.
The
rate of each depolarization/repolarization
that we see has not changed very much.
It
is the generator potential reaching beyond the
effected sites that has been reduced. We know
that this is true because the frequency of
depolarizations has decreased.
Using
100 uM concentrations at the effected nodes of
Ranvier completely eliminated the current
downstream along the pathway.
Notice
also the "washout" diagram (shown here to
illustrate that the nerve returned to
approximately initial condition after tests).
Question:
One of the reasons that three nodes of Ranvier
must be effected is that it can be difficult to
block enough Na channels to prevent depolarization
from occurring at a node of Ranvier. Would
complete elimination of all APs at one
node of Ranvier necessarily be sufficient to
completely block pain from passing to the brain?
Lidocaine in a
living organism diffuses away from the sodium channel. That's
why Lidocaine "wears off" after a few hours. The rate of
diffusion increases with increasing perfusion with blood.
Since most local anaesthetics also cause vasodilation
(relaxing smooth muscles by the same mechanism), it has been
suggested that methods to promote vasoconstriction could
enhance the effects of many local anaesthetics.
Fine Control
of Movements by the CNS
We mentioned in
passing that finely controlled body movements require detailed
control of body musculature. We will be starting the "muscles"
section of this course soon. We will anticipate a little of
the muscle material in today's discussion of muscular control
by neurons.
In our bodies,
movements of the skeleton involve contraction and relaxation
of specific muscles arranged around joints connecting two
bones.
Every muscle
that pulls a joint in one direction is opposed by a muscle
whose contraction leads to the opposite movement (i.e.,
flexors and extensors).
Muscles
consist of bundles of cells called muscle fibers that
shorten when stimulated.
Motor
neurons synapse with muscle fiber cells. Each muscle
fiber has one excitatory synapse from a motor neuron using
acetylcholine as a neurotransmitter.
The magnitude
of the current received from the motor neuron determines how
much each muscle cell contracts and how many muscle cells
are recruited (you'll see how this works, later).
The
magnitude of the current sent through a neuron depends
upon the size of the generator potential that creates it.
This is
true of a current created by a receptor cell that is
sent through an afferent neuron or the current sent from
your brain through an efferent neuron.
Generator
potentials that arise from a receptor cell can be
referred to as "receptor potentials."
Generator
potentials that can be varied (which is really all of
them) in either afferent or efferent neurons are often
referred to as "graded potentials."
A reflex is one
instance in which the receptor potential that occurs in the
afferent neuron is equal to the graded potential that
determines the current in the motor neuron. How do we know
this is true?
Reflexes
We spoke briefly
about reflex arcs in a previous document. Reflexes are
involuntary movements initiated by sensory stimuli.
Recall the
stretch sensor in the knee jerk reflex. Sensory stimuli (like
the stretch sensor) activate interneurons that are part of
simple circuits in brainstem and spinal cord. These circuits
coordinate excitation and inhibition of motorneurons to
initiate simple, highly stereotyped movements such as the
stretch reflex and the pain reflex ("crossed extension
reflex"). The advantage of having reflexes is that they permit
rapid responses when necessary (flight behavior in many
animals). The disadvantage is that movements occur without
input from higher centers and are "unrefined" (i.e, they lack
the capacity for a graded response). Example: when your
daughter tickles you while you are carrying a full cup of
coffee.
The
sensors, which we called "spindle fibers" are also
called "proprioceptors" (sensors that sense themselves
- i.e., the state of the entity that they are part
of).
Courtesy
J.R.
Schiller, Austin Peay State University
AP
frequency is proportional to the speed of
stretch
AP
frequency is proportional to the degree (amount)
of stretch.
Also
important in providing information that relates to
maintenance of posture.
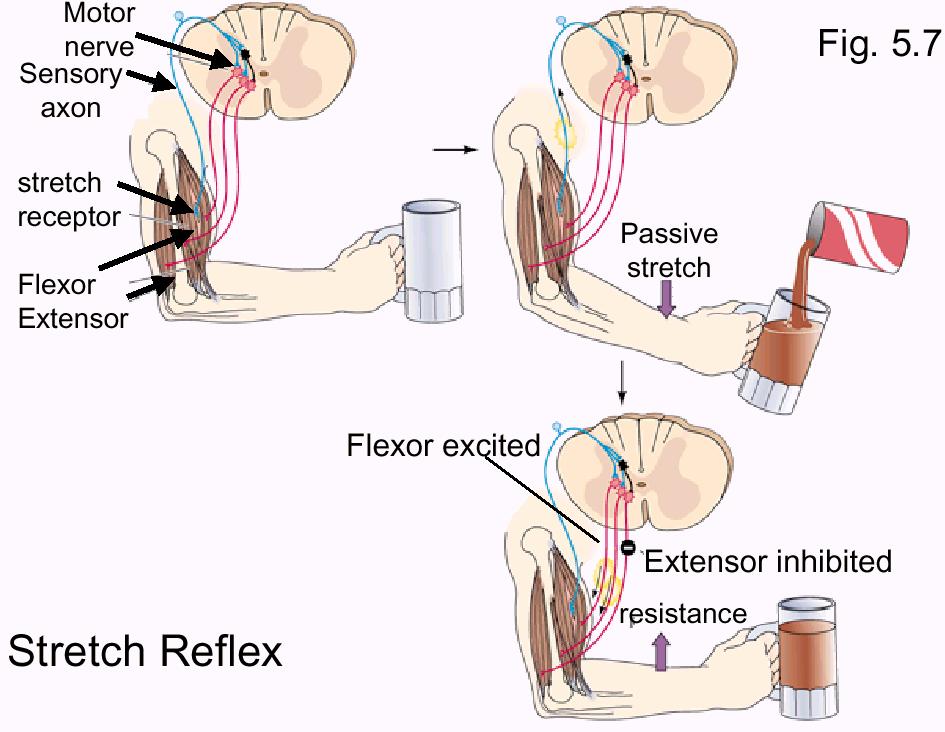
Stretch
Reflex. A stretch reflex is a reflex excitation of
extensors or flexors to maintain fixed body position against a
disruptive force such as gravity (e. g. knee jerk reflex) or
change in load on a muscle (see below). It's purpose is to
work against the applied load (push toward the stimulus) so
that the position of the body is not greatly changed (for
example, as the weight of the mug in the figure below
increases we want the mug to remain in approximately the same
location).
It's initiated
by stimulation of a stretch receptor in a muscle and it
results in increased excitation of synergistic muscles
(muscles that help to reach the end result of the reflex -
i.e., the flexor in the figure below) and inhibition of
antagonist muscles (those muscles that would work against the
purpose of the reflex - i.e., the extensor in the figure
below).
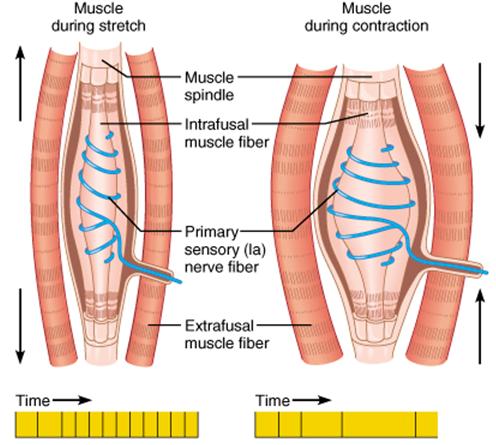
The stretch
reflex shown below is also an example of a graded response.
The amount of flexor contraction appropriate to maintain the
position of a filling mug is different than the amount of
contraction appropriate to maintain position if someone puts a
50 lb sack of cement into your arms. The sack of cement will
result in a greater stretching rate than the filling mug.
Muscle spindles that respond to the stretch rate will produce
AP's at greater frequency in response to the heavier object
and so the reflex response will be a more vigorous muscular
response.
The example
shown above is essentially the same as the "hammer-knee"
example from last reading. Stretching of the propioceptor
results in an AP that arcs back to an excitatory terminal bulb
at the flexor and an inhibitory terminal bulb at the extensor.
The amount of force generated by the reflex is a function of
the rate of stretching of the proprioceptor. A rapid stretch
results in a burst of high frequency AP's that results in a
more vigorous muscular contraction (e.g., being passed a mug
of a drink or a bag of cement results in a different degree of
muscular response).
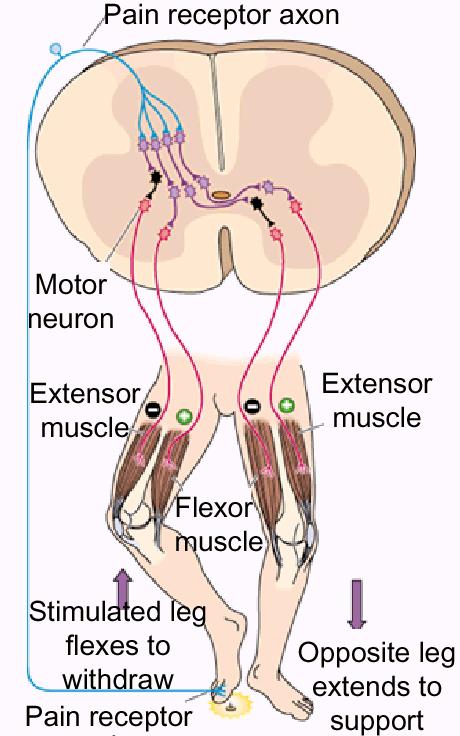
Pain
Reflex (“Crossed Stimulation”). There are times
when you would rather have a reflex that pulled away
from a stimulus rather than pushing toward it.
Stepping on a tack is an example of such a time.
A
pain receptor sends a message that is routed both
to your consciousness ("Ouch!") and to a reflex
circuit.
Using
the figure at right, the reflex excites the motor
neuron to the right side flexor (to pull your foot
back) and inhibits the motor neuron to your right
extensor (to cooperate with the flexor
signal).
At
the same time, it does the opposite to your left
leg: inhibiting the flexor and exciting the
extensor. This helps you to maintain balance while
getting off the tack as expeditiously as
possible.