Other Muscle
Proteins, Force of Contraction, and Muscle Cell
Repair
Introduction
to Today's Reading
The title pretty
much says it all for this reading. Today, we will look at
Some of the
other structural proteins which allow a muscle cell to have
the structure that it does and that also allow it to behave
in ways that we expect (i.e., relax at the end of
contraction) and become constructed properly when they are
created.
We will
look at how we can measure the force generated by a
contracting muscle cell (and in the process, see how the
Frank-Starling mechanism in the heart works).
We will
look more closely at how muscles are constructed during
fetal development and early childhood and, in the process,
see how they repair themselves following injury or exercise
and become stronger.
Review
You should
be able to put the pieces that we have learned so far
together to tell a coherent story.
Structural
components:
Muscle
- Defined by outer covering of connective tissue
(epimyceum)
Fascicle
- Subunit of muscle defined by perimyceum
Motor
unit - muscle fibers (cells) within a fascicle
activated by a single motor neuron.
Muscle fiber - muscle
cell surrounded by endomyceum
Myofibril - cylindrical
structures with the cell defined by
consecutive sarcomeres.
Transverse tubules -
inpouchings of sarcolemma (cell membrane)
Sarcoplasmic reticulum -
Sacks containing Ca
Dihydropyridine
receptors activated by intracellular
current
Ryanodine receptors
activated by DHP act as protein channels
in SR that open to release Ca to
intracellular fluid surrounding
myofibrils.
Ca pump - returns Ca to
SR
Sarcomere - occur end to
end in myofibrils; basic unit of
contraction.
Actin - thin filament
having attachment sites for myosin
heads.
Attachment sites are
covered by tropomyosin when no
contraction going on
Troponin - when Ca
attachment sites are occupied by Ca,
changes conformation, which changes
conformation of tropomyosin, which
uncovers MH attachment sites on actin,
which initiates contraction.
Myosin - thin filaments
Myosin heads - when MH
attachment sites are available, nearby
MH's attach and initiate the
contractal sequance from Quiz 3.
Looking more
closely:
The
arrival of a current in a motor neuron to the
neuromuscular junction causes the sequence of
events you have already learned (VG Ca channels,
myosin motors to move vesicles, snare proteins to
open vesicles and terminal membrane, release of
ACh).
Only
excitation occurs at a NM junction.
ACh
binds to LG Na receptors on the muscle motor end
plate, creating the charge gradient (also called
the generator potential).
On
the muscle side of the neuromuscular junction, the
generator potential initiates current that is
propagated by action potentials.
boosted
by VG Na and repolarized by VG K channels.
The
current moves along the cell membrane
(sarcolemma).
As you learned
studying about neurons, acetylcholinesterase continuously
breaks down the ACh attached to the receptors on the motor end
plate. So, the current through the muscle will cease unless
the current from the neuron to the muscle cell continues.
Current,
propagated by AP's, reaches the cell interior via
the T-tubules.
Current
activates the DiHydroPyradine receptors (voltage
sensors).
The
DHP activates (opens) the Ryodine receptors (Ca
channels in the SR).
Ca
attaches to troponin to shift tropomyosin to
initiate contraction.
Then
there is the myosin-actin contraction sequence
(which you know).
As
long as current continues, contraction continues.
Termination
of current reconforms DHP and closes RyR.
Ca
pump pumps Ca back into SR.
Troponin
and tropomyosin return to their conformation prior
to contraction.
Myosin-Actin
return to original position via elastance of
muscle cell.
Other
Proteins in the Sarcomere
The
"cytoskeleton" is the network of filamentous proteins and
carbohydrate microtubules that extend through the interior of
a cell. Among other things, the cytoskeleton helps to maintain
the shape of the cell, holding the pieces together in their
proper positions.
Actin
and myosin are the components of the cytoskeleton that
get the most attention in muscles.
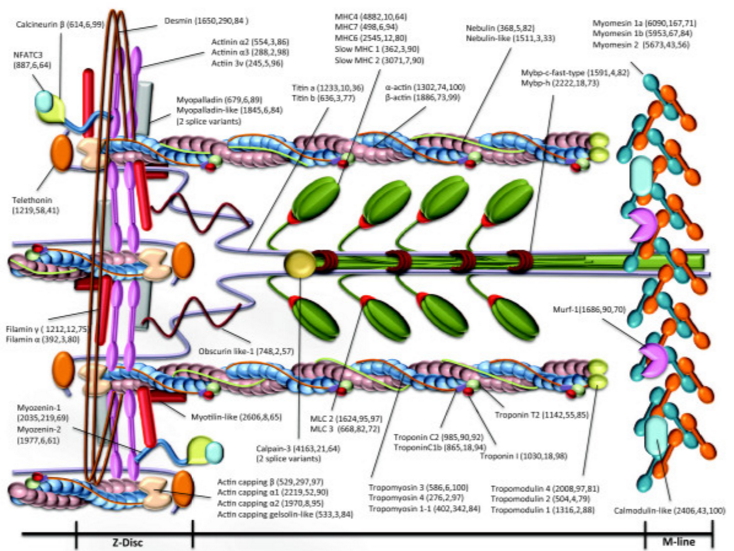
Another
component of a sarcomere is Titin.
"Titin
is the largest polypeptide yet discovered (~3.5 MDa).
Megadalton
- atomic weight of 1 million
Single
molecules span from the Z- to M-line (yellow in the
diagram, below).
These
have two main functions:
In the
thick (myosin) filament part of the myofibril ... the
titin molecule regulates exact myosin assembly by acting
as a giant template or "protein-ruler...
The
remainder of the titin molecule forms an elastic
connection between the end of the thick filaments and
the Z-line.
These
connections give muscle cell its passive tension and they
also keep thick filaments centered between Z-discs.
(Without this there would be force imbalances in the
opposite halves of thick filaments during active
contraction)." Source
The
figure at right demonstrates how titin provides elastic
recovery of muscle length when it has been stretched (by
an opposing muscle) or when it has contracted.
Titin is
not the only protein that gives the sarcomere its
characteristic form.
Nebulin
is shown here.
It
is the foundation protein for actin and
appears
to provide the template for the actin repeating
units.
The "z-disk" is
a combination of proteins that acts as an anchoring point for
the titin and nebulin and mark the ends of the sarcomere.
What you are seeing above is a schematic simplification.
The image at right, which shows half of a sarcomere
(from z-disk on the left to m-line on the right).
Notice that:
the
z-disk is a combination of a number of different
proteins.
The
z-disk anchors both the myosin and actin filaments
on both sides (the z-disk is the place where
sarcomeres are joined).
The
m-line anchors the myosin but does not appear to
anchor the actin.
Desmin
is another important protein.
Desmin
ties the myofibrils together.
It
also anchors the myofibrils to the
sarcolemma.
All of
these proteins provide structural stability (they keep
the components properly positioned relative to each
other) and provide a degree of functionality (template
for building actin, abilility of long axis to relax
into its original length following contraction).
Force of
Contraction: Sarcomere Length and Tension Matters
Force of
contraction in a muscle cell depends on the relative positions
of the myosin and actin filaments.
To get
maximum force when the current arrives, the tension on the
cell (on the myofibrils) must fall within optimum limits.
It's
possible to demonstrate that muscles that start to
contract when they are stretched too far generate less
force of contraction.
It's
possible to demonstrate that the same thig happens when
there is not enough initial tension.
To get
optimum contraction, tension must fall within physiological
limits that normally occur in organisms.
These
experiments described here have been repeated many times.
Muscles
are attached to a stand so that passive force can
be measured.
The
force is only due to the amount of stretch
applied.
The
force is recorded.
Electrical
stimulation is applied to make the muscle
contract.
The
amount of additional force is called active
force.
The
passive force is subtracted from the total force
to determine the force of contraction.
The only thing
that changes in these experiments is the amount of initial
stretch when the muscle is electrically made to contract. This
is really measuring how the initial actin and myosin filament
positions of the sarcomeres affects strength of contraction.
The results of
these tests are summarized in the figure below, right.
There
is an optimum initial degree of stretching (a).
This
corresponds to the "at rest" length of the
sarcomere.
The
MH attachment sites on actin are all close to
the myosin, so when they become available, the
maximum number of MH's will be involved in the
contraction.
If the
initial stretch is greater than normal at rest tension, some
of the MH's and actin attachment sites are no longer
available to each other (b).
As the
number of possible cross bridges decreases, the maximum
possible force of active contraction becomes less.
(when a
MH is attached at an actin attachment site, it is called a
"cross bridge").
If the
stretch becomes great enough, very few MH's can reach actin
attachment sites, and force of active contraction approaches
zero (b, far right).
If the
degree of initial stretch drops below typical at rest
tension, the number of cross bridges also decreases as
"slack" is introduced into their tension.
The actin
and myosin filaments will bend away from each other and
some MH's will not reach actin attachment sites (c).
If the
sarcomeres are actually compressed, some of the MH's will
attach to the wrong part of the actin and push the actin
filament in the wrong direction.
Notice
the actin filament overlap at (d).
So, the
amount of force that a muscle cell can generate depends upon
the proximity of MH's to available actin attachment sites
and normal "at rest" tension for skeletal muscles is pretty
much in the optimal range.
Look at
the left side of the curve.
Do
you recall the Frank-Starling mechanism when we
talked about heart muscle cells?
Does
this figure give you any clues about how the F-S
mechanism works? (it should!).
Skeletal
Muscle Cell Development
Embryology.
Every
mature muscle cell develops from 100 or so
myoblasts (pre-muscle cells) that fuse together in
the fetus. That's the reason skeletal muscle cells
are multinucleated.
Muscle
growth is a result of cellular enlargement &
not cell division.
Muscle
cells have many specialized structures.
In
general, cells having a high degree of
specialization cannot divide and have a difficult
time repairing themselves.
Fetal
Development of Skeletal Muscle Cells
Skeletal muscle
cells are created from the fusion of myoblasts (Greek: myo =
muscle and blast = seed).
Myoblasts
are slightly differentiated stem cells that combine to
create a muscle cell.
This
occurs during the development of the embryo and
continues for a period post birth.
Here is
a more detailed figure.
The
process of fusion begins with the development of
gap junctions (direct cell to cell connections of
the cytosol).
Over
time, the myoblasts physically fuse.
During
this time, thye can depolarize in the presence
of acetylcholine (LG Na channels are present)
and they begin to concentrate calcium.
The
process continues with the fusion of approximately
100 myoblasts per muscle cell.
Myofibrils,
t-tubules, sarcomere etc. development follows
completed fusion.
In this
video, we can watch the above process in action.
This
diagram contains some new terms.
The
"plasmalemma" is a synonym for the cell membrane
(i.e., sarcolemma).
The
basal lamina (and the unlabeled layer above it)
are extracellular layers that separate the cell
membrane from the connective tissue.
The
basal lamina forms as the cell matures.
With
the formation of the basil lamina, it is possible
to see, for the first time, satellite cells.
Satellite
cells divide slowly during post natal
development.
Some
of the cells fuse with the muscle fiber, others
remain separate.
The
article at this link
describes satellite cell motility. Satellite cells
apparently have the ability to migrate about under the
basil lamina!
Satellite
Cells Repair Damaged Muscle Cells
The significance
of satellite cells is that they allow skeletal muscle cells to
be improved and repaired.
Mature
muscle cells cannot divide mitotically, they are just too
complicated.
Muscle
growth (hypertrophy) is a result of cellular enlargement
& not cell division.
Muscle
cells become bigger through the addition of new
myofibrils.
Muscle
cells cannot replace themselves through division, the way
simpler cells do.
This is
typical of complex cells (neurons are like this too).
They also
cannot repair themselves without help from other cells (also
a shared characteristic with neurons).
In general,
cells having a high degree of specialization cannot divide
and have a difficult time repairing themselves.
Neurons don't get
damaged all that frequently. Muscles get damaged all the time.
Despite
the presence of all that protective connective tissue,
the regular contracting and stretching that skeletal
muscles undergo frequently damages the muscle
cells.
The
satellite cells are the mechanism of skeletal muscle
repair.
The
cycle of damage and satellite-cell-mediated repair
is an important mechanism of muscle strengthening.
Here is
an undamaged skeletal muscle cell.
1) Z disk; 2) length of sarcomere; 3) length
of thick filaments; 4) intramuscular triglyceride
droplet; 5) M-line (a cytoskeletal structure to hold
the thick filaments in place. The numerous small
black dots are glycogen molecule complexes.
Here
again is an undamaged skeletal muscle cell (left) and a
sample taken from a runner following a marathon
Here are some
other views of exercise induced muscle cell damage.
In this
electron micrograph, two muscle cell have been split
in two.
Here we
see the process of muscle repair in action.
(A)
Following damage, phagocytic cells (macrophage
"inflammatory cells") clean up the mess by
ingesting cell debris.
(B)
Activated satellite cells divide.
(C)
Daughter cells are myoblasts.
(D)
The myoblasts "recapitulate" embryological
development.
They
go through the same steps that occurred during
fetal development.
They
form myofribrils, sarcolemma, SR, etc.
(E)
the repaired cell.
Here we
see a photographic sequence of the muscle cell repair
process.
There is
some evidence that activated satellite cells from muscle
cells that have been destroyed may replace the dead cell.
I am not
sure about this yet - I found conflicting reports.
When
the cell membrane is disrupted, satellite cells
migrate into the interior of the cell to begin the
repair process.
There
is some evidence that satellite cells may be
activated by exercise without disrupting the
sarcolemma.
Daughter
cells (myoblasts) apparently fuse with the
sarcolemma (much as they fused during fetal
development) and enter the cell to make repairs or
build new myofibrils.
Hypertrophy
is the construction of new myofibrils within a
muscle cell.
The
addition of new myofibrils increases the diameter
of the muscle cell and strengthens the muscle.
Strength
Training (e.g., weight training):
Hypertrophy
(the enlargement of the muscle) is the most
observable result.
Strength
training has both a neural and a mechanical
component.
Neural
adaptation is experienced first and includes:
Faster
motor neuron activation.
Faster
recruitment of muscle cells.
Improved
coordination of the muscle cells that have been recruited.